Exploring Sea Surface Temperature’s Effect on Global Ocean Productivity in Our Changing Climate
{kind=link}

The author aboard UNH's R/V Gulf Challenger just outside of the Judd Gregg Marine Research Complex in New Castle, NH. Photo credit: Sophie Alpert.
Climate change is introducing environmental alterations at a timescale unlike any before and altering ecosystems to a point where resources are becoming severely impacted. The social and ecological value of marine ecosystem stability is vastly dependent on fish biomass, and the fishing industry couples ecosystem health and economic productivity. Constant assessment of the environment where valued species thrive is crucial to ensure sustainable yield of marine species; a failure to detect or act upon observed changes can jeopardize social-ecological systems. Consequently, it is more important than ever to assess the ocean’s stability.
A study conducted for my senior thesis aimed to explore warming and cooling regions of the global ocean and assess the trends in net primary productivity (NPP) with time. NPP represents the difference in the amount of carbon dioxide taken in by phytoplankton during photosynthesis and the amount of carbon dioxide subsequently released via respiration. NPP can be used as a measure of the photosynthetic growth supporting the base of the marine food chain. In the ocean, NPP depends on the overall availability of nutrients that limit phytoplankton biomass. I assessed all of these variables to test the prediction that warmer sea surface waters will result in decreased nutrient availability and marine NPP and that cooler surface waters will result in increased NPP. Decreased NPP rates would result in less CO2 drawdown from the atmosphere by autotrophic phytoplankton, leading to decreased ocean carbon storage. Therefore, more CO2 remains in the atmosphere, further contributing to global warming.
Background
The ocean absorbs heat from the sun, which is then distributed around the global ocean. Heat is a major source of “fuel” for ocean circulation; cold polar waters sink and move equatorward while warm equatorial waters move poleward on what is known as the global conveyor belt. Increased glacial meltwater at the poles disrupts seawater circulation, because less cold water is sinking to depth to drive the circulation process. With a slowing conveyor belt, warm waters are no longer effectively distributed across the global ocean, and we are observing areas of warming but also of cooling sea surface temperature (SST).
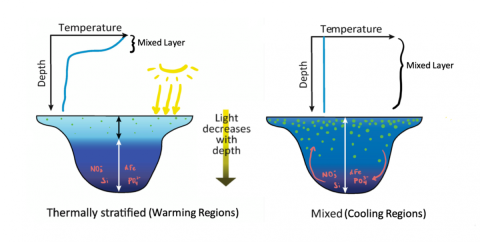
Figure 1a (left) and 1b (right). In figure 1a, solar radiation continuously heats the surface waters, causing thermal stratification. Nutrients produced via photosynthesis accumulate in the lower part of the water column and there is no vertical mixing. In contrast, when water is cooled (figure 1b), it sinks, getting replaced by warmer water continuously to then cool and sink. With this vertical mixing, nutrients that sink have a mechanism to move back up into the surface waters.
It is observed that climate-induced ocean surface warming leads to an increase in thermal stratification. This means there is an increasingly warm layer of surface water where light penetrates, and a barrier forms between that layer and the colder, denser water that sits below, preventing mixing through the water column (Figure 1a). This is thought to prevent the movement of nutrients to the upper water column, which limits phytoplankton growth and photosynthesis rates, thereby having a negative impact on NPP. Thermal stratification is a limiting factor of NPP in warming regions, but in regions where surface waters are cooling, the continuous sinking of cooling waters creates a well-mixed layer of nutrient-rich water at the surface (Figure 1b), thereby having a positive impact on phytoplankton growth and NPP.
Current phytoplankton biogeography suggests that warm tropical and subtropical waters are dominated by smaller species, called picophytoplankton, whereas cool temperate and polar waters are dominated by larger plankton species, including diatoms. Thus, warming of the ocean should shift the ecosystem toward smaller phytoplankton, which exhibit lower NPP rates. The opposite would be expected to occur in cooling regions of the global ocean, where diatom (among other large-cell phytoplankton) biomass concentrations would be expected to increase. Part of my research explored the correlation between different sizes of phytoplankton and the regions in which they flourish to further corroborate evidence for the impact of SST change on NPP rates.
Methods
I used long-term mean SST measurements provided by the National Oceanic and Atmospheric Administration (NOAA) to create an SST anomaly map for 1971–2000. I was interested in exploring the past two decades (2001–2019) of SST change, so I computed the difference between annual SST for each year from 2001–2019 and the long-term (1971–2000) mean SST. By using the calculated long-term mean, I compared the current SST to its historical average, thus indicating whether it has been increasing or decreasing over recent years. To explore whether NPP may be changing and why, I identified four warming regions (North Atlantic, Northwest Pacific, Southeast Pacific, and Southern Ocean) and four cooling regions (Subtropical North Atlantic, Southwest Pacific, Subarctic, and Northeast Pacific)..
I then extracted data specific to each region from the output of the Community Earth System Model (CESM) to quantify both NPP rates within these regions, as well as the following variables to assess reasons for NPP fluctuation:
- inorganic nutrient concentrations, including phosphate, nitrate, silicon, and dissolved iron
- phytoplankton biomass differentiated by size, including diatoms and picophytoplankton
- zooplankton biomass
- nutrient limitations of different phytoplankton types
I used only data from the upper fifty meters of the water column, where primary productivity occurs due to the presence of phytoplankton and light. Data for each of the eight regions was compiled in Microsoft Excel, where the average annual values were recorded and statistically analyzed to diagnose trends in NPP flux.
Results
Preliminary findings suggest a potential correlation between SST and NPP rates. Although my analysis showed only a slight decrease in NPP in regions where SST was rising, this trend may gain significance if assessed over a longer timescale. My analysis also revealed the predicted increase in NPP in regions where SST is declining. This finding supports previous established thought and emphasizes the importance of monitoring SST fluctuation for predicting marine NPP and related support of fisheries.
Inconsistencies in plankton biomass led to results that did not fully support the hypothesis that a future warmer ocean will result in decreased marine NPP. Although diatom biomass decreased in three of the four warming regions and increased in three of the four cooling regions, as one would expect according to current plankton biogeography, and zooplankton (the next level up the food chain) decreased in warming regions, no specific trends in zooplankton biomass were observed within the cooling regions. Further, trends in picophytoplankton lacked significance across both warming and cooling regions assessed in this study. With further research, I hope to expand my timescale of interest in an attempt to diagnose significant trends in plankton biomass
In addition, overall inorganic nutrient concentrations, a key factor in phytoplankton growth, were observed to be decreasing proportionally to NPP rates in three of the four warming regions, further supporting the prediction that higher SST leads to lower NPP. Through my analysis of specific nutrient concentration and limitation trends, I observed phosphate decreasing in essentially all fluctuating temperature regions of the global ocean, whereas nitrate increased in areas of decreasing SST, and vice versa. I also found that iron limitation may act as a buffer on NPP changes. I concluded that further research should target the synergistic effects of nitrate and iron delivery and limitation on diatom growth in fluctuating temperature regions of the global ocean.
I would like to sincerely thank Dr. Robert Letscher for his guidance as a mentor and advisor through my time at UNH. This work would not have been possible without his patience and willingness to train me as a research assistant, especially during this time of COVID-19 uncertainty. It was because of his creativity and flexibility that I was able to complete research outside of the laboratory. This work was partially supported by a student grant from the Hamel Center for Undergraduate Research, whom I would also like to thank for their support in providing the adequate technologies to conduct my research.
Copyright 2021, Jessica Gray